| Dictionnaire médical |
Abréviations M Macr Macro Macu Magn Mal Malacie Maladie Maladie de Malaria Malt Mamm Mamm(o)- Man Manœuvre de Mart Maso Mass Mast Mast(o) Maïe Melæ Mens Menstr Meur Micr Micro Mict Miction Migr Migraine Mili Minéral Miop Mito Mnèm Mném Mnés Moel Mole Moll Mona Mono Morb Morp Mort Moru Mour Muc Muci Muco Mult Musc Muscle Myas Myco Mycé Mydr Myo Myri Myring Myth Myx Myél Méat Méca Méde Médi Médiastin Médu Médull Méga Méio Méla Mélan Méni Méning Ménisc Méno Mér Méro Més Méso Méta Méth Méthode de Métr Métro Mété Météor Môla Môle
| Méiotique | |
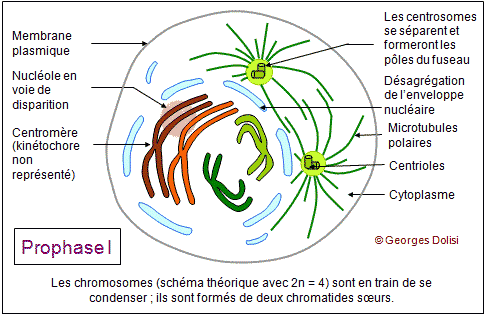
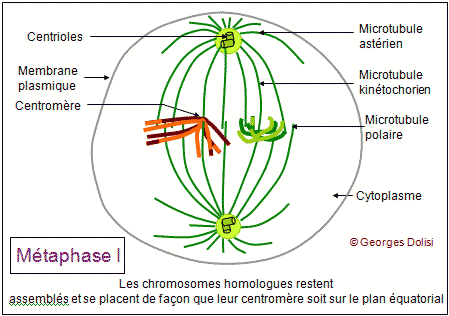
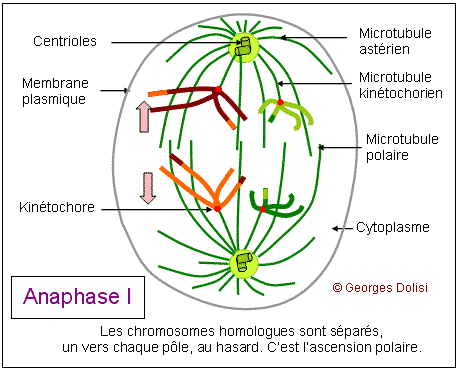
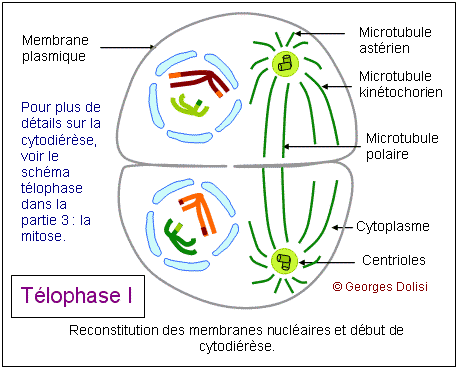
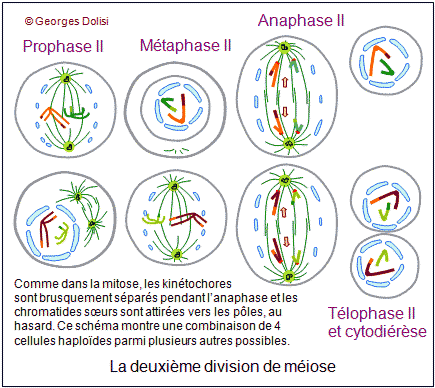
| Cytologie, médecine de la reproduction, génétique Adj. * méiose : du grec meion, meiosis {méio-, mio-}, moindre, décroissance, diminution. Dans notre corps, il y a 2 types de cellules : * Les cellules dites somatiques qui ont 2n (= 46) chromosomes et sont donc diploïdes : elles possèdent 1 exemplaire de chaque chromosome de la mère, soit n = 23, et un exemplaire de chaque chromosome du père, soit n = 23. Si nous transmettions à nos enfants le patrimoine génétique de ces cellules, ils auraient 2n chromosomes de la mère + 2n chromosomes du père, soit 4n, puis 8n à la génération suivante, ce qui est impossible. * C'est la raison pour laquelle nous possédons un deuxième type de cellules, les cellules germinales ou gamètes ou cellules sexuelles, ovocytes et spermatozoïdes, qui sont haploïdes et ne possèdent que n chromosomes, soit un seul exemplaire des 23 chromosomes humains. Ce cette façon, nous transmettons à nos enfants 23 chromosomes maternels dans l'ovule et 23 chromosomes paternels dans le spermatozoïde. Leurs cellules se développeront donc à partir d'un uf ou zygote qui a 2n = 46 chromosomes. Le nombre de chromosomes de l'espèce humaine est ainsi conservé. Il doit donc exister un mécanisme spécial qui permet, à partir de cellules diploïdes, d'obtenir des cellules haploïdes, c'est-à-dire des gamètes. Ce phénomène est en fait une série d'événements appelée la méiose, qui intervient pendant la gamétogenèse et qui permet de passer d'une cellule diploïde à quatre cellules haploïdes chez l'homme, à une cellule haploïde chez la femme. La formation des gamètes ou gamétogenèse sera étudiée en détail au chapitre 5. Ce qui suit n'est donc qu'une étude théorique de la méiose, qui va permettre de répondre à quelques questions fondamentales : - Comment réduire le nombre des chromosomes de 2n à n ? - Si l'on passe d'une cellule diploïde à 4 cellules haploïdes, quels mécanismes qui assurent la présence du même matériel chromosomique dans ces cellules ? - Comment expliquer que pour un même couple, les enfants soient ressemblants mais différents (diversité génétique) ? Première division ou méiose I : réductionnelle exemple théorique avec une cellule de départ à 4 chromosomes (n = 2) Dans cet exemple, 2 chromosomes proviennent de la mère, les 2 autres du père. Un chromosome maternel et le chromosome paternel équivalent forment ce que l'on appelle des chromosomes homologues car ce sont les deux chromosomes d'une même paire. Pendant la phase S, les chromosomes sont dupliqués et les deux chromatides surs d'un même chromosome restent étroitement associées. Dans une division normale (mitose - voir chap. 3), les chromatides surs s'alignent sur le fuseau, leurs fibres kinétochoriennes dirigées vers les pôles opposés. C'est pendant l'anaphase qu'elles se séparent pour devenir des chromosomes individualisés. De cette façon, chaque cellule fille formée par division cellulaire "normale" hérite d'un homologue de chaque chromosome. Dans la méiose, qui aboutit à la formation des gamètes, les cellules ne doivent finalement contenir qu'un seul élément de chaque paire de chromosomes homologues, l'homologue maternel ou l'homologue paternel et donc la moitié seulement du nombre initial de chromosomes. Il y a de ce fait deux divisions successives, mais une seule réplication de l'ADN. 1. La prophase I : De nombreux phénomènes se produisent pendant cette prophase de la première division, ce qui explique qu'elle soit si longue, couvrant à elle seule 90% de la durée totale de la méiose. Les chromosomes dupliqués se condensent progressivement, comme dans la mitose, mais s'unissent ensuite deux par deux, formant des paires d'homologues appelées bivalents. Dans notre exemple, les 4 chromosomes vont donc former 2 paires d'homologues ou 2 bivalents et, pour qu'il n'y ait pas d'erreur au moment de cet appariement, des protéines spécifiques vont assurer la reconnaissance des homologues, ainsi que leur assemblage (un chromosome de la paire n° 3 par ex. ne pourra s'apparier qu'avec l'autre chromosome de la paire n° 3). C'est pendant cette phase qu'un phénomène très particulier se produit : les crossing-over ou échanges réciproques de fragments de chromosomes. Pour bien comprendre le rôle de ces échanges, il faut se rappeler que sur 2 chromosomes homologues, les gènes sont tous placés de la même façon ; ce sont les allèles qui changent. Les crossing-over ne modifient donc pas la succession des gènes, mais uniquement la répartition des allèles. On sait maintenant que ces échanges (coupure puis soudure de fragments d'ADN) sont réalisés grâce à des enzymes de recombinaison très rigoureuses, qui ne permettent que des échanges de régions strictement homologues. Les généticiens ont constaté qu'il y a en moyenne 3 crossing-over (on parle aussi d'enjambements) par bivalent et que ce phénomène est déterminant pour brouiller la constitution génétique de chacun des chromosomes des gamètes, permettant ainsi la recombinaison génétique. Cette phase de la méiose permet l'obtention de chromosomes qualifiés de recombinés. A part les vrais jumeaux, deux individus issus des mêmes parents ne sont pas génétiquement identiques. Cela est dû au fait que, longtemps avant la fusion de l'ovocyte et du spermatozoïde, deux types de brassages génétiques se sont déjà produits pendant la méiose. * Le premier brassage découle de la répartition au hasard des chromosomes homologues maternels et paternels entre les cellules filles pendant l'anaphase I (ou anaphase de première division - voir ci-dessous). Ainsi, avec n = 23 (il y a donc 46 chromosomes sur la plaque équatoriale en métaphase), et sachant que les chromatides d'un même chromosome sont séparées au hasard, il y a déjà 223 possibilités pour les gamètes d'un même individu ! * Un deuxième brassage résulte des phénomènes schématisés ici : les crossing-over ou enjambements qui échangent des segments de chromosomes homologues. De ce fait, le nombre des possibilités pour les gamètes d'un même individus devient gigantesque. Pour en savoir plus sur la prophase I Des changements morphologiques très complexes ont lieu au niveau des chromosomes au moment de leur appariement (synapsis ou syndièse) et de leur séparation (asynapsis ou asyndièse). En fonction de la taille et de la forme des chromosomes, la prophase I a été conventionnellement divisée en 5 stades : * Leptotène : (lepto : du grec leptos, mince, -tène : du grec tenôn, teinô (ten-yo), tendon, tendre, étirer) Pendant le stade leptotène, les chromosomes sont déjà dupliqués, mais pas encore très condensés. C'est parce qu'ils sont encore longs et fins que ce stade est appelé leptotène. * Zygotène : (zygo : du grec zugon [zyg(o)-], joug, paire ; en anatomie, se rapporte à larcade zygomatique (pommette) ou désigne la notion de paire) Le zygotène marque le début de l'appariement deux à deux des chromosomes homologues. Ils sont déjà nettement plus condensés, donc plus courts et plus épais. On est en présence de bivalents. Ils sont attachés par un complexe protéique très particulier qui fonctionne un peu comme une fermeture éclair : le complexe synaptonémal (du grec sunapsis [synapt(o)-, synapse], point de jonction, liaison, union et du grec nêma, nêmatos [nem(o)-, némat(o)-, -nème, -némal], fil, filament). * Pachytène : (pachy : du grec pakhus [pachy-], épais, gros) Pendant le pachytène, la condensation des chromosomes appariés se poursuit et touche à sa fin. C'est aussi pendant cette phase que vont se produire les crossing-over ou enjambements, grâce à l'apparition de nodules de recombinaison qui vont permettre des échanges de chromatine entre le chromosome d'origine maternelle et le chromosome d'origine paternelle. Le complexe synaptonémal est complet. Remarque importante concernant les chromosomes sexuels : Comment l'appariement se fait-il pour les chromosomes sexuels (paire n° 23) qui sont différents, le chromosome X féminin étant nettement plus grand que le chromosome Y masculin ? En réalité, ces deux chromosomes présentent une petite région d'homologie à l'une de leurs extrémités, et cette région est suffisamment grande pour qu'il s'y produise en général un chiasma. C'est ce chiasma qui va permettre à ces deux chromosomes de rester assemblés pendant la métaphase. * Diplotène : (du grec diploos [dipl(o)-], double) Le diplotène marque le début de la séparation, mais la "fermeture éclair" du complexe synaptonémal ne s'ouvre pas intégralement. Les chromosomes homologues restent attachés par les nodules de recombinaison. * Diacinèse : (dia : du préfixe grec dia- signifiant ici séparation, distinction, cinèse : du grec kinêsis, mouvement) La diacinèse marque la fin de la prophase I et la transition avec la métaphase I. Les chromosomes de chaque bivalent s'éloignent l'un de l'autre, surtout au niveau des centromères et forment des figures caractéristiques, comme le montre le schéma du bivalent. 2. La métaphase I : * Les complexes centriolaires se sont déplacés vers les pôles et sont maintenant diamétralement opposés. * Une caractéristique de cette métaphase I est le fait que les chromosomes restent assemblés en bivalents, retenus entre autres par leurs chiasmas. * Tous les bivalents se sont rassemblés sur le plan équatorial, et s'orientent de façon que les kinétochores (il y en a un par chromosome car les 2 kinétochores des chromatides surs ont fusionné) soient dirigés l'un vers un pôle du fuseau, le deuxième vers l'autre pôle. * Les microtubules kinétochoriens s'allongent et rejoignent les pôles, parallèlement aux microtubules du fuseau (ou polaires). 3. L'anaphase I : Elle est fondamentalement différente de la métaphase d'une mitose, car ici ce sont des chromosomes entiers (donc à 2 chromatides) qui vont migrer vers les pôles, alors que dans une mitose, il y a séparation des chromatides, chacune des deux allant vers un pôle ou l'autre de la cellule. Résultat de cette particularité : une nouvelle phase de synthèse pour dupliquer l'ADN n'est pas nécessaire, mais la cellule qui avaient au départ 2n chromosomes à 2 chromatides (donc elle était diploïde) a donné naissance à 2 cellules filles qui n'ont plus que n chromosomes, mais toujours à 2 chromatides, donc à 2 cellules haploïdes. C'est la raison pour laquelle cette première division de méiose est qualifiée de réductionnelle et c'est cette réduction du nombre des chromosomes qui va permettre la conservation du nombre (46) au moment de la fécondation. * Les chromosomes homologues sont tirés chacun vers un pôle différent, au hasard, par les microtubules kinétochoriens qui raccourcissent, en même temps que le fuseau s'allonge. Le brassage intrachromosomique qui a eu lieu pendant la prophase, grâce aux crossing-over et aux échanges de fragments de chromosomes (donc d'ADN) prend maintenant tout son sens car les 2 chromatides surs d'un même chromosome ne sont pas identiques. Le schéma montre une combinaison possible de répartition des chromosomes, parmi un nombre gigantesque. 4. La télophase I : Cette phase est marquée par : * la reconstitution des enveloppes nucléaires autour des bivalents réunis à chaque pôle de la cellule, formant deux noyaux haploïdes, contenant chacun n paires de chromatides. * La division de la cellule en deux cellules filles par cytokinèse. A noter que la télophase I est parfois très incomplète et rapide, enchaînant immédiatement la deuxième division de la méiose. Les chromosomes étant déjà formés de 2 chromatides, une nouvelle synthèse d'ADN n'est pas utile et la prophase II peut débuter sans attendre. Il peut y avoir une brève interphase, pendant laquelle les chromosomes peuvent subir un début de décondensation. Deuxième division ou méiose II : équationnelle suite de notre exemple théorique avec une cellule de départ à 4 chromosomes (n = 2) Cette deuxième partie de la méiose est en tous points comparable à une mitose (voir le chapitre 3) et c'est la raison pour laquelle cette partie se limite à en montrer le résultat et la conséquence au niveau de la diversité génétique. La prophase II est brève. L'enveloppe nucléaire disparaît en même temps que le nouveau fuseau se forme. Pendant la métaphase, les chromosomes se placent sur le plan équatorial du fuseau et les centromères s'accrochent aux kinétochores (un par chromatide). L'anaphase est marquée par l'allongement des cellules et la brusque rupture des kinétochores. Les chromatides surs sont entraînées vers les pôles, au hasard, ce qui augmente la diversité génétique. Pour la télophase et la cytodiérèse, le schéma est volontairement simplifié pour ne montrer que l'une des combinaisons possibles. On voit que les 4 cellules ont toutes un chromosome des 2 paires de départ, mais sont toutes différentes, suite aux échanges d'ADN qui se sont produits pendant les crossing-over de la prophase I. La seule vraie différence entre la méiose II et la mitose est que les chromosomes de ces cellules sexuelles ne sont présents qu'en un seul exemplaire au lieu de deux, ce qui va permettre, lors de la fusion avec l'autre gamète, de conserver le nombre des chromosomes de l'espèce. Pour une application de la méiose, voir la gamétogenèse, c'est-à-dire à la formation des cellules sexuelles, ovogenèse et spermatogenèse. Dans les deux cas, cette gamétogenèse se fait en suivant les différentes étapes de la méiose. L'adjectif mitotique qualifie ce qui se rapporte à la méiose ou ce qui en fait partie.
© Georges Dolisi | |
| < vers le menu principal | Politique de confidentialité | App Privacy Policy | ^ haut de la page |
| © 2006, 2020 Medicopedia | Powered by JIMaroc |