| Dictionnaire médical |
Abrév Abréviations C Cach Caco Cadh Cadu Caec Cail Cal Calc Calcan Calcul Call Camp Campt Cana Canc Cancer Cand Capi Capill Caps Carb Carc Carcin Card Cardi Cardio Carm Cart Cary Carè Caré Cata Cath Cato Caté Caus Cave Cavi Cavo Cavu Cein Cell Cellul Cent Cerv Cervic Cham Chei Chev Chik Chim Chimio Chir Chla Chlo Chlor Chol Chon Chondr Chor Chori Choré Chro Chrom Chroma Chromo Chry Chyl Chym Chéi Chéil Chél Chém Cinq Circ Cirr Cirs Clai Clar Clau Clav Clea Clim Clin Clit Clon Cléid Coag Coar Cobl Cocc Coch Cochlé Coda Code Codo Codé Coeli Col Coli Coll Colo Colp Coma Comp Comé Con Conc Cond Cong Coni Conj Conn Cons Cont Contra Contre Conv Coor Coph Copr Coqu Cord Corn Coro Corp Cort Cortic Cory Cost Coty Cox Coxa Coxo Cran Crani Cras Craw Cres Crev Cric Cris Crow Crue Cruo Crur Crya Crye Cryo Cryp Crypto Crân Créa Créat Crén Crép Cubi Cubit Cura Curc Cure Curi Cut Cyan Cycl Cyph Cyst Cyth Cyto Cli Cur Cæca Cæco Cæcu Céph Céphal Céré Cérébe Cérébr Côlo Côn Cône Côte
| Cycle cellulaire | |
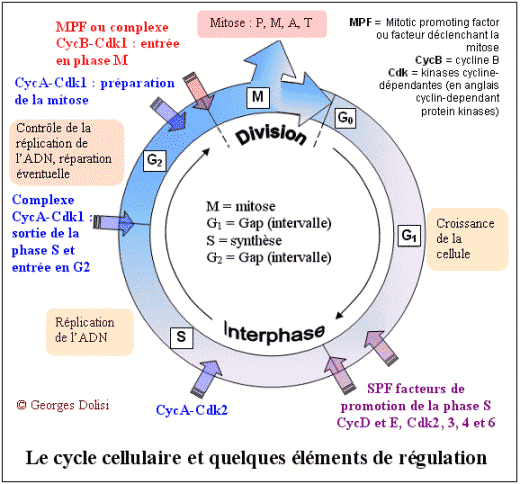
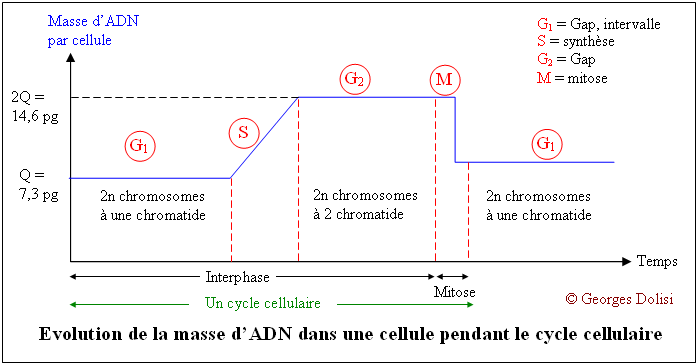
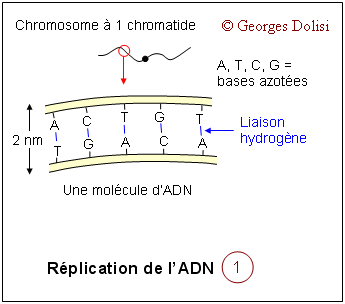
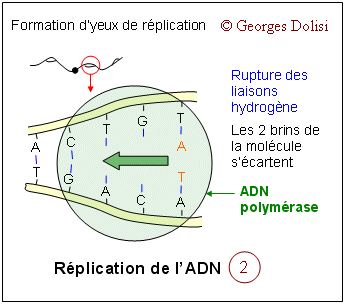
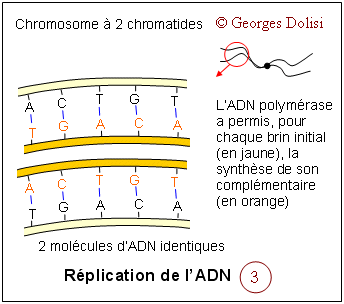
| Cytologie histologie, cancérologie, médecine biologique N. m. * cycle : du grec kuklos {cycl(o)-, -cycle, -cyclique}, cercle, roue ; * cellulaire : du latin cellula, dim. de cella {cellul(o)-, -cellulaire}, chambre ; racine des termes relatifs à la cellule. Le cycle cellulaire (d'une division à la suivante) a une durée très variable selon qu'il s'agisse d'une cellule sanguine, hépatique, épithéliale par exemple, de quelques heures à plus d'un an. La seule constante est la durée de la phase M (la mitose proprement dite) qui est d'environ 1 à 2 heures. Le reste du cycle est qualifié d'interphase : c'est la partie du cycle pendant laquelle la cellule ne se divise pas, est au repos. Le cycle cellulaire est divisé classiquement en quatre phases, dont la plus importante est la mitose, division du noyau puis de la cellule. Phase G1 : G est la première lettre de l'anglais "Gap" ou Gap of time", intervalle. C'est l'intervalle de temps qui s'écoule entre la fin de la mitose et le début de la phase suivante ou synthèse. C'est pendant la phase G1 que la cellule contrôle sa taille et son environnement. A noter que c'est aussi pendant cette phase qu'une cellule qui n'a pas encore commencé à répliquer son ADN peut entrer dans un état quiescent appelé G0, (G zéro) qui peut durer de quelques jours à plusieurs années. Enfin, une cellule qui ne se divise plus reste en phase G1 jusqu'à sa mort. Phase S : ou phase de synthèse, pendant laquelle la cellule réplique son ADN - voir schéma ci-dessous et fabrique les histones qui sont nécessaires pour la confection des nouveaux brins d'ADN. Cette synthèse des histones se fait dans le cytoplasme, puis les nouvelles molécules entrent dans le noyau par les pores nucléaires et se combinent à l'ADN en formant les nucléosomes. On parle de réplication de l'ADN, ou duplication : c'est la copie conforme de l'information génétique, grâce à un complexe de réplication qui contient, entre autres, une enzyme, l'ADN polymérase. Cette réplication est qualifiée de semi conservatrice, car chacune des 2 molécules constituées est formée à partir d'un brin de la molécule initiale. C'est cette synthèse qui fait passer le chromosome de 1 à 2 chromatide, doublant de ce fait sa masse d'ADN. Phase G2 : c'est pendant cette période que la cellule va vérifier que son ADN a été correctement répliqué (elle est donc provisoirement tétraploïde), mais aussi qu'elle va finir sa croissance pour la division de la cellule proprement dite qui suit immédiatement celle du noyau. Phase M : c'est la division de la cellule mère en deux cellules filles strictement identiques (sauf erreurs de réplication). Pour les détails de cette phase : voir ci-dessous. Quelques explications supplémentaires concernant la réplication de l'ADN pendant la phase S : La réplication de l'hélice d'ADN (représentée "à plat" sur les schémas) commence avec la séparation de ses deux chaînes grâce à la rupture des liaisons hydrogène (en bleu sur les schémas). Ces séparations se produisent simultanément à plusieurs endroits de la molécule au niveau des "yeux de réplication". Lorsque la séparation est effective, chaque brin de la molécule mère va servir de matrice pour la synthèse d'un brin complémentaire (à raison d'environ 50 nucléotides par seconde), ce qui va produire finalement 2 hélices filles. L'information génétique est ainsi dupliquée dans son intégralité, de façon que deux doubles hélices complètes d'ADN soient formées, chacune ayant une séquence nucléotidique identique à celle de l'hélice d'ADN parentale qui a servi de matrice. C'est parce que chaque molécule d'ADN fille est constituée d'une chaîne d'origine et d'une chaîne complémentaire synthétisée que l'on a qualifié cette réplication de semi-conservative. Tous ces mécanismes de réplication ne sont possibles que grâce à un complexe multienzymatique, dont l'ADN polymérase et une ligase. C'est en 1957 que la première enzyme de polymérisation nucléotidique a été découverte et nommée ADN polymérase. Elle permet la reconnaissance de chaque nucléotide de l'ADN dont les deux brins ont été séparés, par un nucléotide complémentaire non polymérisé, de façon que les groupes donneurs et accepteurs de liaison hydrogène sur chaque base soient exposés pour l'appariement. Les nucléotides isolés entrants qui conviennent sont alignés pour leur polymérisation par catalyse enzymatique en une nouvelle chaîne d'acide nucléique. Les contrôles du cycle cellulaire Toutes les étapes du cycle cellulaire sont sous la dépendance de 2 familles de protéines interactives (des protéines kinases et des cyclines), constituant un véritable contrôle central qui s'exerce à certains points critiques du cycle (réplication de l'ADN, mitose et cytodiérèse) grâce à des rétrocontrôles émanant des étapes déjà réalisées. Ainsi, une nouvelle phase ne débutera que si la précédente s'est correctement et totalement réalisée. 1. Les protéines kinases Cdk ou protéines kinases cycline-dépendantes (en anglais cyclin-dependant protein kinases) : ce sont des enzymes qui sont capables de transférer un groupement phosphate (phosphorylation) sur des protéines cibles : tyrosine et thréonine. Une protéine phosphorylée (active) par une kinase, peut à tout moment être déphosphorylée par une phosphatase et devient inactive. Ces protéines Cdk vont s'associer à des cyclines pour déclencher les phases du cycle cellulaire. En fait, on connaît actuellement au moins 9 kinases dont Cdk1à Cdk7 qui vont intervenir pendant le cycle cellulaire, en s'associant à des cyclines spécifiques. Cdk 8 et Cdk 9 sont impliquées dans la transcription. A noter que les Cdk étaient antérieurement dénommées CDC (Cell division control) et que CDC2 correspond aujourd'hui à Cdk1. 2. Les cyclines : ce sont des protéines spécialisées qui se lient aux molécules Cdk pour contrôler leur pouvoir de phosphorylation. Cette liaison est indispensable au bon fonctionnement du complexe enzymatique. Les cyclines doivent leur nom au fait qu'elles subissent un cycle de synthèse et de dégradation à chaque cycle cellulaire. Les biologistes distinguent au moins 15 cyclines, dont les cyclines mitotiques (cycA et B) qui se lient aux molécules Cdk1 et Cdk2 pendant la phase G2 et vont permettre le déclenchement de la mitose, et les cyclines G1 (cycD et E) qui vont se lier aux Cdk2, Cdk4 et Cdk6 pendant la phase G1 pour déclencher la phase S (de synthèse). C'est la dégradation des cyclines qui induit la fin des phases S et M. A noter également cette propriété étonnante des cyclines : elles favorisent l'expression des cyclines de la phase suivante et répriment l'expression des cyclines de la phase précédente, tout en favorisant leur dégradation. MPF et SPF : le MPF est le facteur déclenchant la mitose (en anglais Mitotic promoting factor), c'est-à-dire le complexe CycB-Cdk1 (ou CycB-CDC2). Les SPF ou facteurs de promotion de la phase S sont indispensables pour préparer et déclencher la phase S. Ce sont des complexes formés de CycD et CycE, associées à Cdk 2, 4 et 6. 3. Les CKI ou CdkI :cyclin-dependant kinase inhibitor ou protéines inhibitrices des kinases cycline-dépendantes. Ces protéines capables d'inhiber les Cdk en s'associant avec elles ou avec le complexe Cyc-Cdk interviennent en interdisant le passage à la phase suivante. On comprend mieux le rôle fondamental de ces CKI dans la régulation ou la suppression des tumeurs. Elles sont connues sous les noms de : * p21 : si des lésions de l'ADN sont apparues pendant le cycle cellulaire et notamment pendant la phase S, une protéine p53 va activer p21 qui est un inhibiteur de Cdk. Il en résulte un arrêt du cycle cellulaire en G1, avec soit réparation de l'ADN, soit apoptose (mort cellulaire). * p27 : comme la précédente, cette protéine empêche le passage de la phase G1 à la phase S. * p57 : agissant aussi en bloquent le passage G1 --> S, on a constaté que, chez les humains, une expression aberrante de la protéine p57 est associée à divers néoplasmes, ainsi qu'au syndrome de BECKWITH-WIEDEMANN.
© Georges Dolisi | |
| < vers le menu principal | Politique de confidentialité | App Privacy Policy | ^ haut de la page |
| © 2006, 2020 Medicopedia | Powered by JIMaroc |